Двойное оплодотворение у растений
Двойное оплодотворение у растений имеет большое биологическое значение. Оно было открыто Навашиным в 1898 г. Далее рассмотрим подробнее, как происходит двойное оплодотворение у растений.
Биологическое значение
Процесс двойного оплодотворения способствует активному развитию питательной ткани. В связи с этим семяпочка не запасает вещества впрок. Это, в свою очередь, объясняет ее быстрое развитие.
Схема двойного оплодотворения
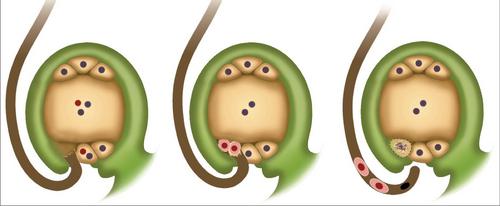
Коротко явление можно описать следующим образом. Двойное оплодотворение у покрытосеменных растений состоит в проникновении в завязь двух спермиев. Один сливается с яйцеклеткой. Это способствует началу развития диплоидного зародыша. Второй спермий соединяется с центральной клеткой. В результате формируется триплоидный элемент. Из этой клетки появляется эндосперм. Он является питательным материалом для развивающегося зародыша.
Развитие пыльцевой трубки
Двойное оплодотворение у покрытосеменных начинается после образования гаплоидного сильно редуцированного поколения. Оно представлено гаметофитами. Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.

Формирование клеток: общие сведения
Процесс двойного оплодотворения цветковых растений осуществляется особыми половыми клетками. Их формирование происходит в два этапа. Первая стадия называется спорогенез, вторая – гематогенез. В случае образования мужских клеток эти этапы именуются микроспорогенез и микрогематогенез. При образовании женских половых элементов приставка меняется на "мега" (или "макро"). Спорогенез основывается на мейозе. Это процесс формирования гаплоидных элементов. Мейозу, так же как и у представителей фауны, предшествует размножение клеток посредством митотических делений.
Образование спермиев
Первичное формирование мужских половых элементов осуществляется в особой ткани пыльника. Она называется археспориальной. В ней в результате митозов происходит формирование многочисленных эелементов – материнских клеток пыльцы. Они и вступают затем в мейоз. Вследствие двух мейотических делений образуется 4 гаплоидные микроспоры. Некоторое время они лежат рядом, формируя тетрады. После этого происходит их распад на пыльцевые зерна - отдельные микроспоры. Каждый из образованных элементов начинает покрываться двумя оболочками: внешней (экзина) и внутренней (интина). Затем начинается следующий этап – микрогаметогенез. Он, в свою очередь, состоит из двух митотических последовательных делений. После первого формируется две клетки: генеративная и вегетативная. Впоследствии первая проходит еще одно деление. В результате образуется две мужские клетки – спермии.

Макроспорогенез и мегаспорогенез
В тканях семяпочки начинает обособляться один или несколько археспориальных элементов. Они начинают усиленно расти. Вследствие такой активности они становятся значительно крупнее остальных клеток, окружающих их в семяпочке. Каждый археспориальный элемент один, два или более раз подвергается делению митозом. В некоторых случаях клетка может сразу трансформироваться в материнскую. Внутри нее происходит мейоз. В результате него формируется 4 гаплоидные клетки. Как правило, самая крупная из них начинает развиваться, превращаясь в зародышевый мешок. Три оставшиеся постепенно дегенерируют. На данном этапе макроспорогенез завершается, начинается макрогематогенез. В ходе него происходят митотические деления (у большей части покрытосеменных их три). Цитокинез не сопровождает митозы. В результате трех делений формируется зародышевый мешок с восемью ядрами. Они впоследствии обосабливаются в самостоятельные клетки. Эти элементы распределяются определенным образом по зародышевому мешку. Одна из обособленных клеток, которая, собственно, является яйцеклеткой, совместно с двумя другими – синергидами, занимает место у микропиле, в которое осуществляется проникновение спермиев. В этом процессе синергиды исполняют очень значимую роль. В них содержатся ферменты, которые способствуют растворению оболочек на пыльцевых трубках. В противоположной стороне зародышевого мешка располагаются другие три клетки. Они именуются антиподами. С помощью этих элементов происходит передача из семяпочки питательных веществ в зародышевый мешок. Оставшиеся две клетки располагаются в центральной части. Зачастую они сливаются. В результате их соединения формируется диплоидная центральная клетка. После того как произойдет двойное оплодотворение, и в завязь проникнут спермии, один из них, как выше сказано было, сольется с яйцеклеткой.

Особенности пыльцевой трубки
Двойное оплодотворение сопровождается взаимодействием ее с тканями спорофита. Оно достаточно специфично. Этот процесс регулируется активностью химических соединений. Установлено, что если пыльцу промыть в дистиллированной воде, она потеряет способность к прорастанию. Если же полученный раствор сконцентрировать, а затем ее обработать, то она снова станет полноценной. Развитие пыльцевой трубки после прорастания контролируют ткани пестика. К примеру, у хлопчатника ее рост до яйцеклетки занимает порядка 12-18-ти часов. Однако уже спустя 6 часов вполне можно определить, к какой именно семяпочке будет направляться пыльцевая трубка. Это понятно потому, что в ней начинается разрушение синергиды. В настоящее время не установлено, как растение может направить развитие трубки в нужном направлении и каким образом о приближении узнает синергида.
"Запрет" на самоопыление
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. "Запрет" на самоопыление проявляется в том, что спорофит "идентифицирует" собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же – короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца – мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.
Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев "самонесовместимость" пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, - s1s2, а вырабатывающего пыльцу - s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.

В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
Похожие статьи
- Гамета - это половая клетка: строение, виды, функции
- Можно ли забеременеть до овуляции: ответ гинеколога
- При каком размере плодного яйца виден эмбрион? Размеры эмбриона по неделям
- Гаметогенез – это процесс формирования половых клеток
- Бесполое размножение и его виды
- Доказательство эволюции - эмбриологические стадии развития животных
- Что такое зародыш: строение, процесс формирования и развития