Дрейф генов - фактор эволюции: причина, значение, примеры
Наряду с естественным отбором существует и другой фактор, способный оказать влияние на повышение содержания мутантного гена. В ряде случаев он даже может вытеснить нормальный аллеломорф. Называется это явление "дрейф генов в популяции". Рассмотрим более подробно, что собой представляет этот процесс и каковы его последствия.
Общие сведения
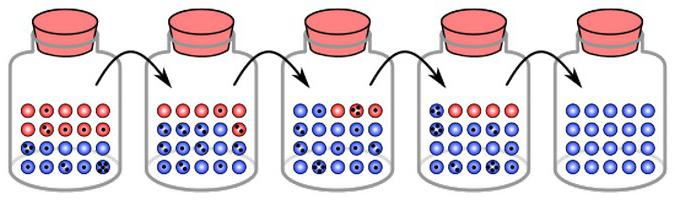
Дрейф генов, примеры которого будут приведены в статье далее, представляет собой определенные изменения, которые фиксируются от поколения к поколению. Считается, что у этого явления существуют свои собственные механизмы. Некоторые исследователи обеспокоены тем, что в генофонде многих, если не всех, наций в настоящий момент довольно быстро увеличивается объем появляющихся аномальных генов. Они определяют наследственную патологию, формируют предпосылки для развития множества других заболеваний. Считается также, что патоморфоз (изменение признаков) разных болезней, в том числе и болезней психического характера, обусловливает именно дрейф генов. Явление, о котором идет речь, происходит стремительными темпами. В итоге ряд психических расстройств принимает неизвестные формы, становятся неузнаваемыми при сопоставлении с их описанием в классических изданиях. Вместе с этим существенные изменения отмечаются и непосредственно в самой структуре психиатрической заболеваемости. Так, дрейф генов стирает некоторые формы шизофрении, встречавшиеся ранее. Вместо них появляются такие патологии, которые с трудом можно определить по современным классификаторам.
Теория Райта
Случайный дрейф генов изучался с помощью математических моделей. Используя этот принцип, Райт вывел теорию. Он считал, что решающее значение дрейфа генов при постоянных условиях отмечается в небольших группах. Они становятся гомозиготными, и изменчивость уменьшается. Райт также полагал, что вследствие изменений в группах способны сформироваться негативные наследственные признаки. В результате этого вся популяция может погибнуть, не внеся вклад в развитие вида. Вместе с этим большую роль во многих группах играет отбор. В этой связи генетическая изменчивость внутри популяции вновь будет несущественной. Постепенно группа хорошо приспособится к окружающим условиям. Однако последующие эволюционные изменения будут зависеть от возникновения благоприятных мутаций. Эти процессы проходят достаточно медленно. В этой связи и эволюция больших популяций не отличается высокой скоростью. В группах промежуточной величины отмечается повышенная изменчивость. При этом образование новых выгодных генов происходит случайно, что, в свою очередь, ускоряет эволюцию.
Выводы Райта
Когда из популяции теряется один аллель, появиться он может вследствие определенной мутации. Но если вид разделяется на несколько групп, в одной из которых отсутствует один элемент, в другой – другой, то ген может мигрировать оттуда, где он есть, туда, где его нет. Таким образом сохранится изменчивость. Учитывая это, Райт сделал вывод, что быстрее развитие будет происходить у тех видов, которые разделены на многочисленные различные по величине популяции. При этом между ними возможна и некоторая миграция. Райт был согласен с тем, что естественный отбор играет весьма существенную роль. Однако вместе с этим результатом эволюции является дрейф генов. Он определяет продолжительные изменения внутри вида. Кроме этого, Райт полагал, что множество отличительных признаков, возникших посредством дрейфа, были безразличны, а в ряде случаев даже вредны для жизнеспособности организмов.

Споры исследователей
По поводу теории Райта существовало несколько мнений. К примеру, Добжанский полагал, что бессмысленно ставить вопрос о том, какой из факторов более значим – естественный отбор либо генетический дрейф. Объяснял он это их взаимодействием. По сути, вероятны следующие ситуации:
- В случае если в развитии тех или иных видов отбор занимает главенствующую позицию, будет отмечаться либо направленное изменение генных частот, либо стабильное состояние. Последнее будет определяться окружающими условиями.
- Если в течение продолжительного периода более существенным будет дрейф генов, то направленные изменения не будут обусловлены природной средой. При этом неблагоприятные признаки, даже возникшие в небольшом количестве, способны достаточно широко распространиться в группе.
Следует, однако, отметить, что непосредственно сам процесс изменений, как и причина дрейфа генов, сегодня изучены недостаточно. В этой связи единого и конкретного мнения о данном явлении в науке не существует.
Дрейф генов – фактор эволюции
Благодаря изменениям отмечается смена частот аллелей. Это будет происходить до того момента, пока они не достигнут состояния равновесия. То есть дрейф генов – изоляция одного элемента и фиксация другого. В разных группах такие изменения происходят независимо. В этой связи итоги генетического дрейфа в разных популяциях различны. В конечном счете в одних фиксируется какой-то один набор элементов, в других – другой. Дрейф генов, таким образом, с одной стороны, приводит к уменьшению разнообразия. Однако вместе с этим он обусловливает и различия между группами, дивергенции по некоторым признакам. Это, в свою очередь, может выступать в качестве основы для видообразования.
Соотношение влияния
В процессе развития генетический дрейф взаимодействует с прочими факторами. Прежде всего, взаимосвязь устанавливается с естественным отбором. Соотношение вкладов данных факторов находится в зависимости от ряда обстоятельств. В первую очередь его определяет интенсивность отбора. Вторым обстоятельством является численность группы. Так, если интенсивность и численность высоки, случайные процессы обладают ничтожно малым влиянием на динамику генетических частот. При этом в небольших группах при несущественных различиях в приспособленности влияние изменений несравнимо больше. В таких случаях возможна фиксация менее адаптивного аллеля, при том что более адаптивный будет потерян.
Последствия изменений
Одним из основных результатов генетического дрейфа выступает обеднение разнообразия внутри группы. Это происходит за счет утраты одних аллелей и фиксации других. Процесс мутации, в свою очередь, напротив, способствует обогащению внутри популяций генетического разнообразия. За счет мутирования утерянный аллель может возникать снова и снова. В связи с тем что генетический дрейф представляет собой направленный процесс, одновременно со снижением внутрипопуляционного разнообразия увеличивается различие между локальными группами. Противодействует этому явлению миграция. Так, если в одной популяции фиксируется аллель "А", а в другой – "а", то внутри этих групп снова появляется разнообразие.

Конечный итог
Результатом генетического дрейфа станет полное устранение одного аллеля и закрепление другого. Чем чаще элемент встречается в группе, тем будет выше вероятность его фиксации. Как показывают некоторые расчеты, возможность закрепления равна частоте аллеля в популяции.
Мутации
Они происходят в среднем с частотой 10-5 на ген на гамету на поколение. Все аллели, которые обнаруживаются в группах, когда-то возникли вследствие мутации. Чем меньше популяция, тем ниже вероятность того, что каждое поколение будет иметь хотя бы одну особь - носителя новой мутации. При численности в сто тысяч у каждой группы потомков с вероятностью, приближенной к единице, найдется мутантный аллель. Однако его частота в популяции, а также возможность его закрепления будет достаточно низкой. Вероятность, что эта же мутация появится в том же поколении хотя бы у одной особи при численности 10, ничтожно мала. Однако если она все же произойдет в данной популяции, то частота мутантного аллеля (1 на 20 аллелей), а также шансы на его фиксацию будут относительно высокими. В больших популяциях возникновение нового элемента происходит относительно быстро. При этом его закрепление проходит медленно. Малочисленные популяции, напротив, мутацию ожидают долго. Но после ее возникновения закрепление проходит быстро. Из этого можно сделать следующий вывод: шанс на фиксацию нейтральных аллелей находится в зависимости только от частоты мутационного возникновения. При этом численность популяции на этот процесс не влияет.
Молекулярные часы
В связи с тем, что частоты появления нейтральных мутаций у разных видов приблизительно одинаковы, скорость закрепления должна быть также примерно равной. Из этого следует, что количество изменений, скопившихся в одном гене, должно соотноситься со временем независимой эволюции данных видов. Другими словами, чем длительнее будет период с момента отделения двух видов из одного предкового, тем больше они различают мутационных замен. Этот принцип лежит в основе метода молекулярных эволюционных часов. Так определяется время, которое прошло с момента, когда предыдущие поколения различных систематических групп начали развиваться самостоятельно, не завися друг от друга.
Исследования Поллинга и Цукуркендла
Эти два американских ученых выявили, что число различий в аминокислотной последовательности в цитохроме и гемоглобине у тех или иных видов млекопитающих тем выше, чем раньше произошло расхождение их эволюционных путей. Впоследствии данная закономерность была подтверждена большим объемом экспериментальных данных. Материал включал в себя десятки различных генов и несколько сотен видов животных, микроорганизмов и растений. Выяснилось, что ход молекулярных часов осуществляется с постоянной скоростью. Это открытие, собственно говоря, подтверждается рассматриваемой теорией. Калибровка часов производится отдельно для каждого гена. Это обусловлено тем, что частота появления нейтральных мутаций у них различна. Для этого проводится оценка количества замен, скопившихся в определенном гене у таксонов. Их время дивергенции надежно установлено с помощью палеонтологических данных. После того как будут откалиброваны молекулярные часы, их можно использовать далее. В частности, с их помощью легко измерить время, в течение которого произошла дивергенция (расхождение) между различными таксонами. Это возможно даже в том случае, если общий их предок еще не выявлен в палеонтологической летописи.
Похожие статьи
- Генетика человека: понятие, основы, методы изучения
- Пример стабилизирующего отбора, признаки и последствия
- Что такое микроэволюция? В чем различие макро- и микроэволюции?
- Виды эволюции: история развития и определение
- Доказательство эволюции - эмбриологические стадии развития животных
- Что такое комбинативная изменчивость?
- Инбридинг - что такое в биологии?